
 Tweet
Tweet
لماذا يوجد تفسيرات كثيرة لتطور دماغ الرئيسيات؟البرفسور روبن دنبار
Why are there so many explanations for primate brain evolution?
لقد حظي السؤال عن سبب تطور أدمغة الرئيسيات بشكل غير عادي باهتمام كبير ، مع العديد من المقترحات البديلة المدعومة جميعها بالأدلة. نقوم بمراجعة الفرضيات الرئيسية والافتراضات التي يقدمونها والأدلة المؤيدة والمعارضة لها.
مع الأخذ في الاعتبار حقيقة أن كل فرضية لديها دليل تجريبي قوي لدعمها كنقطة انطلاق لنا ، فإننا نجادل بأن الفرضيات يتم تفسيرها بشكل أفضل من حيث إطار الأسباب التطورية (عوامل الاختيار) ، والعواقب (النوافذ التطورية للفرص) والقيود (عادةً القيود الفسيولوجية التي تتطلب حلًا إذا كان على العقول الكبيرة أن تتطور).
يجب النظر إلى تفسيرات تطور الدماغ في الطيور والثدييات بشكل عام ، والرئيسيات على وجه الخصوص ، على خلفية التحديات التي ينطوي عليها تطور مجموعات اجتماعية منسقة ومتماسكة ومترابطة تتطلب سلوكيات اجتماعية جديدة لحلها ، جنبًا إلى جنب مع المتخصصين. الإدراك والركائز العصبية التي تدعم هذا.
من القضايا الحاسمة ، ولكن غالبًا ما يتم تجاهلها ، حقيقة أن تطور الأدمغة الكبيرة تطلب التغلب على قيود الميزانية النشطة والفسيولوجية والوقتية.
في بعض الحالات ، انعكس هذا في تطور "البحث الذكي" والذكاء التقني ، ولكن في كثير من الحالات تطلبت تطوير الكفاءات السلوكية (مثل تشكيل الائتلاف) التي تتطلب مهارات معرفية جديدة.
قد تكون هذه كلها مدعومة بنموذج عام من الإدراك يمكن استخدامه في العديد من السياقات المختلفة. هذه المقالة جزء من الموضوع تحت عنوان "المحددات الفسيولوجية للسلوك الاجتماعي عند الحيوانات
مقدمة
سيطر على تطور الرئيسيات ، مثل أي شيء آخر ، أدمغة كبيرة بشكل غير عادي [1].
على مدى العقود الأربعة الماضية ، تم اقتراح العديد من التفسيرات لتطور الأدمغة الكبيرة.
بشكل عام ، تنقسم هذه التفسيرات إلى أربعة موضوعات رئيسية ، لكل منها العديد من الفرضيات الفرعية الخاصة بها:
التفسيرات الجينية (تمتلك الرئيسيات أدمغة كبيرة لأن طفرة جينية معينة تسمح لها بتنمية أدمغة كبيرة) ،
وتفسيرات النمو (الرئيسيات لديها أدمغة كبيرة لأنها ممتدة. تسمح فترات الاستثمار الأبوي لهم بتنمية أدمغة كبيرة) ،
والتفسيرات البيئية (طورت الرئيسيات أدمغة كبيرة من أجل التعامل مع الظروف البيئية الصعبة) ،
والتفسيرات الاجتماعية (هناك شيء معقد جوهريًا حول المجتمع الرئيسى يتطلب دماغًا كبيرًا).
في كثير من النواحي ، كانت المشكلة الرئيسية المرتبطة بفهم سبب تطور الأدمغة الكبيرة هي حقيقة وجود إحراج للثراء: هناك أدلة تجريبية تدعم كل فرضية. ومع ذلك ، فإن حقيقة أنه يمكن تقديم الدليل لصالح تفسيرات مختلفة جدًا من الناحية المفاهيمية ، وغير متوافقة بشكل متبادل ، يجب أن تنبهنا إلى حقيقة أن شيئًا ما غير صحيح. إما أن تكون هناك قضايا إحصائية مربكة مثل أننا نكافح لفصل العلاقات السببية بين مجموعة من السمات شديدة الترابط أو أن هناك قضايا مفاهيمية ناشئة عن بناء تفسيرات بديلة.
نجادل هنا بأن الجهود المبذولة لتحديد التفسيرات الصحيحة لتطور حجم الدماغ قد تعثرت بسبب أربع قضايا رئيسية.
المشكلة الاولى ، والأكثر شيوعًا بلا شك ، هو الفشل في التمييز بين مستويات التفسير المختلفة (المعنى [2]): غالبًا ما يفترض المؤلفون ضمنيًا أن الدليل على فرضية واحدة يقوض جميع التفسيرات الأخرى على الرغم من أن الفرضيات قيد الاختبار قد تكون في الواقع مكملة وضرورية بنفس القدر.
كانت المشكلة الثانية هي أن العديد من الدراسات لا تزال تقدم دليلًا على فرضية واحدة فقط ، وتفشل في الاختبار بشكل مناسب بين البدائل .
كانت المشكلة الثالثة هي الفشل في التحديد الدقيق لكيفية تأثير حجم الدماغ على الإدراك (على سبيل المثال ، ما هي بالضبط جوانب سلوك الرئيسيات التي تتطلب الكثير من الناحية المعرفية) وكيف يرتبط هذا ببيولوجيا الأعصاب الأساسية [3،4].
كانت المشكلة الرابعة هي الميل إلى تفضيل الفرضيات التي تنطبق فقط على مجموعة فرعية من الأنواع ، حتى داخل الرئيسيات.
قد تكون التفسيرات التي تنطبق فقط على حالات خاصة صحيحة ، لكنها لا يمكن أن تكون تفسيرات عامة.
لتسليط الضوء على كيف أعاقت هذه القضايا فهمنا لتطور دماغ الرئيسيات ، نقوم بتقييم كل من التفسيرات الرئيسية وتحديد الافتراضات والقيود المحتملة التي تكمن وراء كل منها على حدة.
من خلال القيام بذلك ، نقوم بتطوير إطار عمل يوضح التفسيرات المختلفة ضمن نموذج توضيحي واحد.
يجب أن نجادل بأن نقطة ارتكاز مهمة في هذا هي التكاليف النشطة لتطوير وصيانة الأدمغة الكبيرة والمجموعات الكبيرة: يتم تجاهل هذه دائمًا. الاختبار الذي يجب أن نطبقه على أي تفسير محتمل هو أنه يمكن أن يشرح ست نتائج تجريبية رئيسية:
(1) أن الرئيسيات لديها أدمغة أكبر بالنسبة لحجم أجسامها من جميع الحيوانات الأخرى [1] ؛
(2) أن بعض الرئيسيات لديها أدمغة أكبر من الرئيسيات الأخرى [1] ؛
(3) أن هناك علاقة كمية قوية بشكل ملحوظ بين حجم الدماغ (وخاصة القشرة المخية الحديثة) وحجم المجموعة في الرئيسيات (ولكن ليس الثدييات أو الطيور الأخرى) [5،6] ؛
(4)أن الرئيسيات لديها شكل غريب من أشكال التواصل الاجتماعي التي تبدو مختلفة تمامًا عن تلك الخاصة بالثدييات الأخرى [7-12] ، وهو ما ينعكس في حقيقة أن مجتمعات الرئيسيات منظمة للغاية من حيث الشبكات (في حين أن مجتمعات الثدييات الأخرى و الطيور ليست) [13-16] ؛
(5) أن الزواج الأحادي في الطيور والثدييات يرتبط بحجم دماغ أكبر من المتوسط بالنسبة لأوامرهم [7،8] ؛
(6) أن بعض (وليس كل) أنواع الرئيسيات تظهر كفاءات فنية جديدة [17 ، 18].
على الرغم من أن العديد من التحليلات تفعل ذلك ، يجب ألا نتجاهل العلاقة بين حجم المجموعة وحجم الدماغ في الرئيسيات ، لأن العيش في مجموعات مكلف للغاية للحيوانات.
هذا هو الحال لثلاثة أسباب.
أولاً ، يؤدي زيادة حجم المجموعة إلى زيادة المنافسة بشكل لا مفر منه ويؤدي إلى التكاليف من حيث الوقت اللازم للبحث عن الطعام والسفر ، وفي الرئيسيات ، الترابط الاجتماعي ، مما يضع ضغطًا كبيرًا على قدرة الحيوانات على البقاء في موطن معين [19].
ثانيًا ، الكائن الحي هو نظام بيولوجي متكامل وأي تغيير في جزء واحد من النظام سيكون له حتمًا تداعيات على أجزاء أخرى. زيادة كتلة الدماغ ، على سبيل المثال ، تفرض متطلبات إضافية من الطاقة والمغذيات ، والتي تتطلب بدورها مزيدًا من الاستثمار في البحث عن الطعام.
يمكن أن تؤدي ضغوط الطاقة هذه إلى قيام الحيوانات بالاستثمار في البحث عن الطعام الأكثر خطورة والأكثر استهلاكا للوقت ، مما قد يعرض نفسها لمخاطر افتراس أعلى.
تزيد هذه الضغوط من ضغط ميزانيات الوقت وقدرتها على الاستثمار في الأنشطة الأساسية الأخرى ، بما في ذلك الترابط الاجتماعي.
الوقت هو قضية رئيسية بالنسبة للحيوانات ، وخاصة الرئيسيات ، ولا ينبغي التقليل من أهميته.
ثالثًا ، تفرض الحياة الجماعية تكاليف فسيولوجية كبيرة على الإناث ، على وجه الخصوص ، بسبب تأثير الإجهاد الاجتماعي على الغدد الصماء في نظام الحيض ، وبالتالي العقم.
باختصار ، لا يمكن ببساطة رفض حجم المجموعة على أنه منتج ثانوي غير رسمي لامتلاك دماغ كبير - إنه جزء أساسي من القصة بسبب حلقات التغذية الراجعة في العلاقات بين هذه المتغيرات.
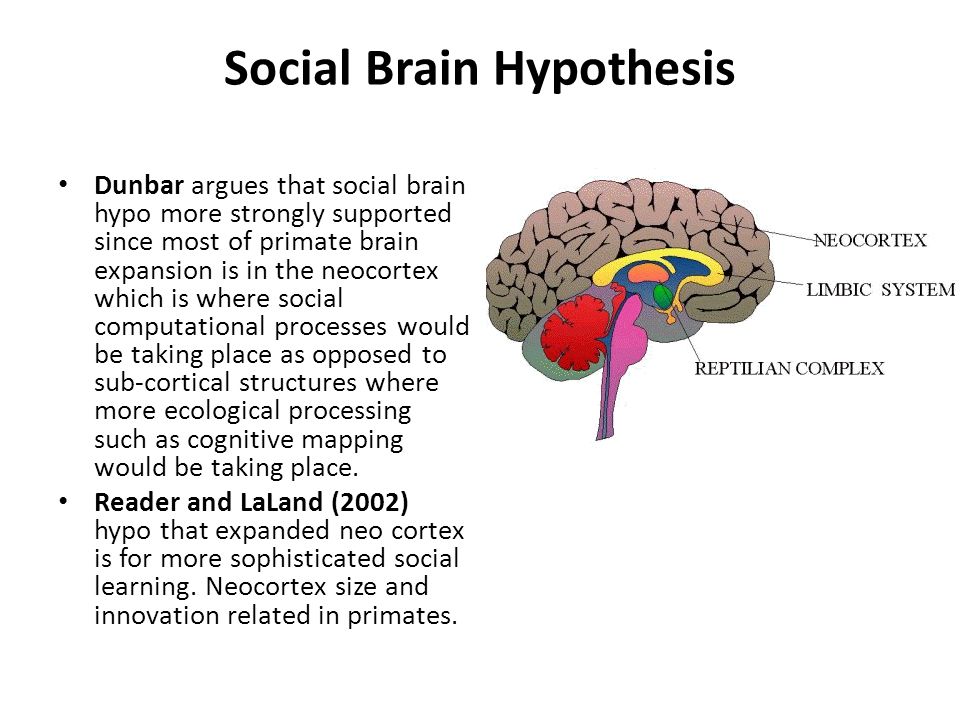
لاحظ أنه ، للأغراض الحالية ، سنقوم بتأطير مناقشتنا بشكل أساسي من حيث حجم الدماغ ، بدلاً من تحديد مناطق معينة من الدماغ. على الرغم من أن أدمغة الرئيسيات الكبيرة هي نتيجة الزيادة الهائلة في حجم القشرة المخية الحديثة [20،21] ، فإن معظم العلاقات السلوكية التي نناقشها ترتبط إلى حد ما بأي مؤشر لحجم الدماغ تقريبًا ، ولا نرغب في أن نكون جانبًا. -تتبعه في مناقشات غير ضرورية حول ما إذا كانت بعض مناطق الدماغ أكثر أهمية من غيرها.
نلاحظ أدناه أن المقاييس المتعددة لهندسة الدماغ تدعم استنتاجات مماثلة.
تمت مناقشة الأسباب التي تجعل الحجم المطلق للدماغ هو أفضل مقياس لاستخدامه كبديل للقدرة المعرفية لدى الرئيسيات من قبل مؤلفين آخرين [17 ، 18].
هناك نقطة أخيرة تستحق التأكيد عليها: تستند جميع الدراسات حول هذا الموضوع تقريبًا إلى أدلة ارتباطية.
لطالما عانت الفرضيات التطورية من العيب المتمثل في أننا لا نستطيع بسهولة اختبار السببية في ما هو ضمنيًا فرضيات سببية.
من وقت لآخر ، يتم إجراء التجارب ، ولكن التحليلات الموضوعية المبنية على هذه تكون دائمًا مرتبطة بالضرورة (حجم الدماغ يرتبط بأداء المهمة) [18] ومع ذلك ، هناك مناهج بديلة يمكن استخدامها الآن ، وسنناقش أن هذه الأساليب تحتاج إلى مزيد من الاهتمام

لماذا وكيف تتطور العقول الكبيرة
لتوفير إطار عمل ، نلخص الفرضيات المختلفة التي تم اقتراحها ، وآثارها الوظيفية ، في الشكل 1.
نحن نبني هذا باعتباره شجرة قرارات يتم فيها تضييق الخيارات تدريجياً من اليسار إلى اليمين.
في الزاوية اليسرى العلوية ، نسرد عددًا من العوامل التي غالبًا ما تم اقتراحها كفرضيات حسنة النية لتطور الدماغ ولكنها في الواقع قيود على حجم الدماغ بدلاً من التفسيرات الوظيفية.
من الضروري عدم الخلط بين التفسيرات الآلية والوظيفية [2].
تعمل القيود التطورية عادة من خلال فرص تنموية محدودة (ما هي الاحتمالات المتاحة في ضوء مجموعة الصفات الحالية [43]).
فيما يتعلق بتطور الدماغ ، ركزت القيود التنموية على سمات تاريخ الحياة [22،25-27] والتفسيرات العصبية [20،21،44] ، ولكن التفسيرات الجينية مؤخرًا (وخاصة تلك الجينات المرتبطة بالتطور المتسارع داخل الإنسان النسب [28–33،45]) انضموا إلى هذه المجموعة.
يمكن أن تنطوي القيود التطورية أيضًا على التكاليف الفسيولوجية لتنمية السمات والحفاظ عليها [23]. الأدمغة مكلفة للغاية [46-48] ، وهذه القيود تمثل بعض التكاليف التي يجب أن تكون الحيوانات قادرة على استيعابها من أجل تطوير أدمغة كبيرة إذا كان لديهم سبب مقنع للقيام بذلك.
تقليديا ، هذه تشمل معدل التمثيل الغذائي [24،49] ، ومتطلبات الطاقة أو الغذائية [22،24،46،50،51].
تطوير حلول للتغلب على هذه القيود ضرورية ، ولكنها ليست كافية ، تفسيرات لتطور الأدمغة الكبيرة.
جميع التفسيرات المتبقية في الشكل 1 هي ، من حيث المبدأ ، تفسيرات وظيفية (أي أنها تقدم ادعاءات حول قوى الاختيار التي ربما تكون قد دفعت تطور الدماغ).
إنهم ينقسمون بشكل طبيعي على بعدين: أولاً ، من خلال ما إذا كانت الحيوانات تحل مشاكل الحد من اللياقة البدنية بشكل فردي (عن طريق التعلم عن طريق التجربة والخطأ أو البصيرة) أو اجتماعيًا (وجود العديد من الأفراد ضروري بشكل صريح حتى يكون الحل فعالًا) ، ثم ، ثانيًا ، من خلال ما إذا كان عامل تحديد اللياقة البدنية مباشرًا (شراء الطعام أو الرفقاء) أو غير مباشر (على سبيل المثال ، ضمان التنسيق الجماعي لإدارة تهديد خارجي للبقاء أو الخصوبة).
الفرضيات الآلية
تركز هذه الفرضيات بشكل أساسي على متطلبات البحث عن الطعام وتفترض ضمنيًا (ولكن ليس بشكل صريح تقريبًا) أن البحث عن الطعام هو أهم قيد على لياقة الحيوان.
في الواقع ، هذا هو الوضع الافتراضي لعلماء البيئة. في التحليلات المبكرة ، كان يُفترض أن المقتصد [25] يتطلب معرفيًا أكثر من الأوراق ، وقد يكون الأمر كذلك: الفاكهة أقل قابلية للتنبؤ بها في الزمان والمكان من الأوراق.
ومع ذلك ، فإن التحليلات المقارنة للتطور لا تجد أي علاقة بين درجة المقتصد الغذائي وحجم الدماغ عند التحكم في حجم المجموعة الاجتماعية عبر الثدييات [6،34،52] - على الرغم من أن الحقيقة الأخيرة قد تكون الهبة الحاسمة من حيث أنها قد تشير إلى أن التغيير من النظام الغذائي ضروري عندما تشارك مجموعات كبيرة بسبب تأثير حجم المجموعة على الطاقة [19].
والأهم من ذلك ، ربما ، لكي يكون للبحث الذكي عن الطعام أي تأثير للتفسير ، من الضروري إظهار أن الرئيسيات تفعل شيئًا مختلفًا عن غير الرئيسيات - وإلا لماذا تحتاج إلى أدمغة أكبر من الثدييات الأخرى؟ لهذا السبب ، ركزت الدراسات الحديثة على ابتكارات البحث عن العلف ، بما في ذلك اكتشاف واستغلال الأطعمة الجديدة [52] أو وسائل جديدة للوصول إلى الأطعمة [34].
أظهر عدد من التحليلات أن ابتكارات البحث عن العلف ترتبط بحجم الدماغ في كل من الطيور والرئيسيات [17،53 –55] ، وهذه العلاقة بدورها مرتبطة بقدرة الأنواع على البقاء في الموائل الصعبة (الطيور [56-60] ، الرئيسيات [61،62] وأشباه البشر [62-66]).
يكمن ضعف هذا الادعاء في أن معظم الأصناف لا تُظهر في الواقع الكثير من البحث الذكي أو الابتكار التقني ، على الرغم من التباين في حجم الدماغ عبر الأنواع. الحقيقة الحاسمة هي أنه في الرئيسيات ، تبدو العلاقة أكثر من مرحلة انتقالية: معظم الأنواع لا تظهر أي ابتكارات على الإطلاق وقليل منها تظهر الكثير [17].
بالنظر إلى هذا ، يبدو أنه تمدد نقطة للادعاء بأن ما هو في الواقع انقسام في الابتكار هو المسؤول عن التغيير الكمي في حجم الدماغ عبر النظام بأكمله. قد يكون التفسير البديل الواضح هو أن البحث الذكي هو نتاج ثانوي لاكتساب دماغ بحجم أدنى معين (أي اختراق سقف زجاجي لتطور الدماغ).
ما الدليل الذي يشير إلى أن اتخاذ القرار البيئي أو التقني له عواقب فعلية على اللياقة؟ قامت دراسة واحدة فقط بتقييم هذا بشكل مباشر: أظهر ألتمان [67،68] أن إناث قرد البابون التي كانت أكثر قدرة على التوافق مع النظام الغذائي الأمثل (من حيث تناول الطاقة والبروتين) مع بقاء الصغار على قيد الحياة لفترة أطول ، وكان لديهم وظائف إنجابية أطول وأنجبوا نسلًا أكثر ( سواء تم تصنيفها على أنها إجمالي عدد المواليد أو عدد البقاء على قيد الحياة حتى 12 شهرًا).
على الرغم من أن حجم العينة صغير (ن ¼ 6 إناث) ، فإن النتائج خطية ومقنعة بشكل ملحوظ. توفر الأدمغة الكبيرة بالتأكيد القدرة على الانخراط في حل المشكلات بطريقة فعالة عن طريق التجربة والخطأ أو التعلم الثاقب لمحاكمة واحدة [69].
هناك قضية أكثر أهمية تتعلق بافتراض أن الغذاء هو ، أو موازنات الطاقة عن طريق الامتداد ، العامل الأساسي الذي يؤثر على لياقة الحيوان ، إما لأن جميع التأثيرات الخارجية الأخرى تافهة بالمقارنة أو لأن البحث عن الطعام هو العامل الوحيد الذي يمكن للحيوان التحكم به فعليًا من خلال سلوكه .
في الواقع ، بالنسبة للثدييات بشكل عام ، والرئيسيات على وجه الخصوص ، يعتبر الافتراس أيضًا أحد الاعتبارات الرئيسية [70-72] وله تأثير أكبر بكثير على التوزيعات البيوجغرافية للأنواع [19،73] - وهذا على نطاق واسع عبر الثدييات والطيور بشكل عام .
فرديًا وجماعيًا ، يبدو أن الفرضيات الآلية تفشل كتفسير عام لأنها لا توضح سبب احتياج الرئيسيات إلى أدمغة أكبر من أي شخص آخر ، ولماذا تختلف أدمغة الرئيسيات كثيرًا في الحجم بين الأنواع ، ولماذا يجب أن يكون لدى الرئيسيات مجموعات اجتماعية مرتبطة ، ولماذا يجب أن يكون حجم المجموعة يرتبط بحجم الدماغ أو لماذا تحتاج الأنواع المتزاوجة إلى مثل هذه العقول الكبيرة. بشكل عام ، إذن ، قد يتم شرح مهارات البحث عن الطعام بشكل أفضل كحل للقيود المفروضة على نمو الأدمغة الكبيرة والحفاظ عليها عندما تكون مطلوبة لسبب آخر.
يتبع في المشاركة الثانية

Why are there so many explanations for primate brain evolution?
لقد حظي السؤال عن سبب تطور أدمغة الرئيسيات بشكل غير عادي باهتمام كبير ، مع العديد من المقترحات البديلة المدعومة جميعها بالأدلة. نقوم بمراجعة الفرضيات الرئيسية والافتراضات التي يقدمونها والأدلة المؤيدة والمعارضة لها.
مع الأخذ في الاعتبار حقيقة أن كل فرضية لديها دليل تجريبي قوي لدعمها كنقطة انطلاق لنا ، فإننا نجادل بأن الفرضيات يتم تفسيرها بشكل أفضل من حيث إطار الأسباب التطورية (عوامل الاختيار) ، والعواقب (النوافذ التطورية للفرص) والقيود (عادةً القيود الفسيولوجية التي تتطلب حلًا إذا كان على العقول الكبيرة أن تتطور).
يجب النظر إلى تفسيرات تطور الدماغ في الطيور والثدييات بشكل عام ، والرئيسيات على وجه الخصوص ، على خلفية التحديات التي ينطوي عليها تطور مجموعات اجتماعية منسقة ومتماسكة ومترابطة تتطلب سلوكيات اجتماعية جديدة لحلها ، جنبًا إلى جنب مع المتخصصين. الإدراك والركائز العصبية التي تدعم هذا.
من القضايا الحاسمة ، ولكن غالبًا ما يتم تجاهلها ، حقيقة أن تطور الأدمغة الكبيرة تطلب التغلب على قيود الميزانية النشطة والفسيولوجية والوقتية.
في بعض الحالات ، انعكس هذا في تطور "البحث الذكي" والذكاء التقني ، ولكن في كثير من الحالات تطلبت تطوير الكفاءات السلوكية (مثل تشكيل الائتلاف) التي تتطلب مهارات معرفية جديدة.
قد تكون هذه كلها مدعومة بنموذج عام من الإدراك يمكن استخدامه في العديد من السياقات المختلفة. هذه المقالة جزء من الموضوع تحت عنوان "المحددات الفسيولوجية للسلوك الاجتماعي عند الحيوانات
مقدمة
سيطر على تطور الرئيسيات ، مثل أي شيء آخر ، أدمغة كبيرة بشكل غير عادي [1].
على مدى العقود الأربعة الماضية ، تم اقتراح العديد من التفسيرات لتطور الأدمغة الكبيرة.
بشكل عام ، تنقسم هذه التفسيرات إلى أربعة موضوعات رئيسية ، لكل منها العديد من الفرضيات الفرعية الخاصة بها:
التفسيرات الجينية (تمتلك الرئيسيات أدمغة كبيرة لأن طفرة جينية معينة تسمح لها بتنمية أدمغة كبيرة) ،
وتفسيرات النمو (الرئيسيات لديها أدمغة كبيرة لأنها ممتدة. تسمح فترات الاستثمار الأبوي لهم بتنمية أدمغة كبيرة) ،
والتفسيرات البيئية (طورت الرئيسيات أدمغة كبيرة من أجل التعامل مع الظروف البيئية الصعبة) ،
والتفسيرات الاجتماعية (هناك شيء معقد جوهريًا حول المجتمع الرئيسى يتطلب دماغًا كبيرًا).
في كثير من النواحي ، كانت المشكلة الرئيسية المرتبطة بفهم سبب تطور الأدمغة الكبيرة هي حقيقة وجود إحراج للثراء: هناك أدلة تجريبية تدعم كل فرضية. ومع ذلك ، فإن حقيقة أنه يمكن تقديم الدليل لصالح تفسيرات مختلفة جدًا من الناحية المفاهيمية ، وغير متوافقة بشكل متبادل ، يجب أن تنبهنا إلى حقيقة أن شيئًا ما غير صحيح. إما أن تكون هناك قضايا إحصائية مربكة مثل أننا نكافح لفصل العلاقات السببية بين مجموعة من السمات شديدة الترابط أو أن هناك قضايا مفاهيمية ناشئة عن بناء تفسيرات بديلة.
نجادل هنا بأن الجهود المبذولة لتحديد التفسيرات الصحيحة لتطور حجم الدماغ قد تعثرت بسبب أربع قضايا رئيسية.
المشكلة الاولى ، والأكثر شيوعًا بلا شك ، هو الفشل في التمييز بين مستويات التفسير المختلفة (المعنى [2]): غالبًا ما يفترض المؤلفون ضمنيًا أن الدليل على فرضية واحدة يقوض جميع التفسيرات الأخرى على الرغم من أن الفرضيات قيد الاختبار قد تكون في الواقع مكملة وضرورية بنفس القدر.
كانت المشكلة الثانية هي أن العديد من الدراسات لا تزال تقدم دليلًا على فرضية واحدة فقط ، وتفشل في الاختبار بشكل مناسب بين البدائل .
كانت المشكلة الثالثة هي الفشل في التحديد الدقيق لكيفية تأثير حجم الدماغ على الإدراك (على سبيل المثال ، ما هي بالضبط جوانب سلوك الرئيسيات التي تتطلب الكثير من الناحية المعرفية) وكيف يرتبط هذا ببيولوجيا الأعصاب الأساسية [3،4].
كانت المشكلة الرابعة هي الميل إلى تفضيل الفرضيات التي تنطبق فقط على مجموعة فرعية من الأنواع ، حتى داخل الرئيسيات.
قد تكون التفسيرات التي تنطبق فقط على حالات خاصة صحيحة ، لكنها لا يمكن أن تكون تفسيرات عامة.
لتسليط الضوء على كيف أعاقت هذه القضايا فهمنا لتطور دماغ الرئيسيات ، نقوم بتقييم كل من التفسيرات الرئيسية وتحديد الافتراضات والقيود المحتملة التي تكمن وراء كل منها على حدة.
من خلال القيام بذلك ، نقوم بتطوير إطار عمل يوضح التفسيرات المختلفة ضمن نموذج توضيحي واحد.
يجب أن نجادل بأن نقطة ارتكاز مهمة في هذا هي التكاليف النشطة لتطوير وصيانة الأدمغة الكبيرة والمجموعات الكبيرة: يتم تجاهل هذه دائمًا. الاختبار الذي يجب أن نطبقه على أي تفسير محتمل هو أنه يمكن أن يشرح ست نتائج تجريبية رئيسية:
(1) أن الرئيسيات لديها أدمغة أكبر بالنسبة لحجم أجسامها من جميع الحيوانات الأخرى [1] ؛
(2) أن بعض الرئيسيات لديها أدمغة أكبر من الرئيسيات الأخرى [1] ؛
(3) أن هناك علاقة كمية قوية بشكل ملحوظ بين حجم الدماغ (وخاصة القشرة المخية الحديثة) وحجم المجموعة في الرئيسيات (ولكن ليس الثدييات أو الطيور الأخرى) [5،6] ؛
(4)أن الرئيسيات لديها شكل غريب من أشكال التواصل الاجتماعي التي تبدو مختلفة تمامًا عن تلك الخاصة بالثدييات الأخرى [7-12] ، وهو ما ينعكس في حقيقة أن مجتمعات الرئيسيات منظمة للغاية من حيث الشبكات (في حين أن مجتمعات الثدييات الأخرى و الطيور ليست) [13-16] ؛
(5) أن الزواج الأحادي في الطيور والثدييات يرتبط بحجم دماغ أكبر من المتوسط بالنسبة لأوامرهم [7،8] ؛
(6) أن بعض (وليس كل) أنواع الرئيسيات تظهر كفاءات فنية جديدة [17 ، 18].
على الرغم من أن العديد من التحليلات تفعل ذلك ، يجب ألا نتجاهل العلاقة بين حجم المجموعة وحجم الدماغ في الرئيسيات ، لأن العيش في مجموعات مكلف للغاية للحيوانات.
هذا هو الحال لثلاثة أسباب.
أولاً ، يؤدي زيادة حجم المجموعة إلى زيادة المنافسة بشكل لا مفر منه ويؤدي إلى التكاليف من حيث الوقت اللازم للبحث عن الطعام والسفر ، وفي الرئيسيات ، الترابط الاجتماعي ، مما يضع ضغطًا كبيرًا على قدرة الحيوانات على البقاء في موطن معين [19].
ثانيًا ، الكائن الحي هو نظام بيولوجي متكامل وأي تغيير في جزء واحد من النظام سيكون له حتمًا تداعيات على أجزاء أخرى. زيادة كتلة الدماغ ، على سبيل المثال ، تفرض متطلبات إضافية من الطاقة والمغذيات ، والتي تتطلب بدورها مزيدًا من الاستثمار في البحث عن الطعام.
يمكن أن تؤدي ضغوط الطاقة هذه إلى قيام الحيوانات بالاستثمار في البحث عن الطعام الأكثر خطورة والأكثر استهلاكا للوقت ، مما قد يعرض نفسها لمخاطر افتراس أعلى.
تزيد هذه الضغوط من ضغط ميزانيات الوقت وقدرتها على الاستثمار في الأنشطة الأساسية الأخرى ، بما في ذلك الترابط الاجتماعي.
الوقت هو قضية رئيسية بالنسبة للحيوانات ، وخاصة الرئيسيات ، ولا ينبغي التقليل من أهميته.
ثالثًا ، تفرض الحياة الجماعية تكاليف فسيولوجية كبيرة على الإناث ، على وجه الخصوص ، بسبب تأثير الإجهاد الاجتماعي على الغدد الصماء في نظام الحيض ، وبالتالي العقم.
باختصار ، لا يمكن ببساطة رفض حجم المجموعة على أنه منتج ثانوي غير رسمي لامتلاك دماغ كبير - إنه جزء أساسي من القصة بسبب حلقات التغذية الراجعة في العلاقات بين هذه المتغيرات.
لاحظ أنه ، للأغراض الحالية ، سنقوم بتأطير مناقشتنا بشكل أساسي من حيث حجم الدماغ ، بدلاً من تحديد مناطق معينة من الدماغ. على الرغم من أن أدمغة الرئيسيات الكبيرة هي نتيجة الزيادة الهائلة في حجم القشرة المخية الحديثة [20،21] ، فإن معظم العلاقات السلوكية التي نناقشها ترتبط إلى حد ما بأي مؤشر لحجم الدماغ تقريبًا ، ولا نرغب في أن نكون جانبًا. -تتبعه في مناقشات غير ضرورية حول ما إذا كانت بعض مناطق الدماغ أكثر أهمية من غيرها.
نلاحظ أدناه أن المقاييس المتعددة لهندسة الدماغ تدعم استنتاجات مماثلة.
تمت مناقشة الأسباب التي تجعل الحجم المطلق للدماغ هو أفضل مقياس لاستخدامه كبديل للقدرة المعرفية لدى الرئيسيات من قبل مؤلفين آخرين [17 ، 18].
هناك نقطة أخيرة تستحق التأكيد عليها: تستند جميع الدراسات حول هذا الموضوع تقريبًا إلى أدلة ارتباطية.
لطالما عانت الفرضيات التطورية من العيب المتمثل في أننا لا نستطيع بسهولة اختبار السببية في ما هو ضمنيًا فرضيات سببية.
من وقت لآخر ، يتم إجراء التجارب ، ولكن التحليلات الموضوعية المبنية على هذه تكون دائمًا مرتبطة بالضرورة (حجم الدماغ يرتبط بأداء المهمة) [18] ومع ذلك ، هناك مناهج بديلة يمكن استخدامها الآن ، وسنناقش أن هذه الأساليب تحتاج إلى مزيد من الاهتمام
لماذا وكيف تتطور العقول الكبيرة
لتوفير إطار عمل ، نلخص الفرضيات المختلفة التي تم اقتراحها ، وآثارها الوظيفية ، في الشكل 1.
نحن نبني هذا باعتباره شجرة قرارات يتم فيها تضييق الخيارات تدريجياً من اليسار إلى اليمين.
في الزاوية اليسرى العلوية ، نسرد عددًا من العوامل التي غالبًا ما تم اقتراحها كفرضيات حسنة النية لتطور الدماغ ولكنها في الواقع قيود على حجم الدماغ بدلاً من التفسيرات الوظيفية.
من الضروري عدم الخلط بين التفسيرات الآلية والوظيفية [2].
تعمل القيود التطورية عادة من خلال فرص تنموية محدودة (ما هي الاحتمالات المتاحة في ضوء مجموعة الصفات الحالية [43]).
فيما يتعلق بتطور الدماغ ، ركزت القيود التنموية على سمات تاريخ الحياة [22،25-27] والتفسيرات العصبية [20،21،44] ، ولكن التفسيرات الجينية مؤخرًا (وخاصة تلك الجينات المرتبطة بالتطور المتسارع داخل الإنسان النسب [28–33،45]) انضموا إلى هذه المجموعة.
يمكن أن تنطوي القيود التطورية أيضًا على التكاليف الفسيولوجية لتنمية السمات والحفاظ عليها [23]. الأدمغة مكلفة للغاية [46-48] ، وهذه القيود تمثل بعض التكاليف التي يجب أن تكون الحيوانات قادرة على استيعابها من أجل تطوير أدمغة كبيرة إذا كان لديهم سبب مقنع للقيام بذلك.
تقليديا ، هذه تشمل معدل التمثيل الغذائي [24،49] ، ومتطلبات الطاقة أو الغذائية [22،24،46،50،51].
تطوير حلول للتغلب على هذه القيود ضرورية ، ولكنها ليست كافية ، تفسيرات لتطور الأدمغة الكبيرة.
جميع التفسيرات المتبقية في الشكل 1 هي ، من حيث المبدأ ، تفسيرات وظيفية (أي أنها تقدم ادعاءات حول قوى الاختيار التي ربما تكون قد دفعت تطور الدماغ).
إنهم ينقسمون بشكل طبيعي على بعدين: أولاً ، من خلال ما إذا كانت الحيوانات تحل مشاكل الحد من اللياقة البدنية بشكل فردي (عن طريق التعلم عن طريق التجربة والخطأ أو البصيرة) أو اجتماعيًا (وجود العديد من الأفراد ضروري بشكل صريح حتى يكون الحل فعالًا) ، ثم ، ثانيًا ، من خلال ما إذا كان عامل تحديد اللياقة البدنية مباشرًا (شراء الطعام أو الرفقاء) أو غير مباشر (على سبيل المثال ، ضمان التنسيق الجماعي لإدارة تهديد خارجي للبقاء أو الخصوبة).
الفرضيات الآلية
تركز هذه الفرضيات بشكل أساسي على متطلبات البحث عن الطعام وتفترض ضمنيًا (ولكن ليس بشكل صريح تقريبًا) أن البحث عن الطعام هو أهم قيد على لياقة الحيوان.
في الواقع ، هذا هو الوضع الافتراضي لعلماء البيئة. في التحليلات المبكرة ، كان يُفترض أن المقتصد [25] يتطلب معرفيًا أكثر من الأوراق ، وقد يكون الأمر كذلك: الفاكهة أقل قابلية للتنبؤ بها في الزمان والمكان من الأوراق.
ومع ذلك ، فإن التحليلات المقارنة للتطور لا تجد أي علاقة بين درجة المقتصد الغذائي وحجم الدماغ عند التحكم في حجم المجموعة الاجتماعية عبر الثدييات [6،34،52] - على الرغم من أن الحقيقة الأخيرة قد تكون الهبة الحاسمة من حيث أنها قد تشير إلى أن التغيير من النظام الغذائي ضروري عندما تشارك مجموعات كبيرة بسبب تأثير حجم المجموعة على الطاقة [19].
والأهم من ذلك ، ربما ، لكي يكون للبحث الذكي عن الطعام أي تأثير للتفسير ، من الضروري إظهار أن الرئيسيات تفعل شيئًا مختلفًا عن غير الرئيسيات - وإلا لماذا تحتاج إلى أدمغة أكبر من الثدييات الأخرى؟ لهذا السبب ، ركزت الدراسات الحديثة على ابتكارات البحث عن العلف ، بما في ذلك اكتشاف واستغلال الأطعمة الجديدة [52] أو وسائل جديدة للوصول إلى الأطعمة [34].
أظهر عدد من التحليلات أن ابتكارات البحث عن العلف ترتبط بحجم الدماغ في كل من الطيور والرئيسيات [17،53 –55] ، وهذه العلاقة بدورها مرتبطة بقدرة الأنواع على البقاء في الموائل الصعبة (الطيور [56-60] ، الرئيسيات [61،62] وأشباه البشر [62-66]).
يكمن ضعف هذا الادعاء في أن معظم الأصناف لا تُظهر في الواقع الكثير من البحث الذكي أو الابتكار التقني ، على الرغم من التباين في حجم الدماغ عبر الأنواع. الحقيقة الحاسمة هي أنه في الرئيسيات ، تبدو العلاقة أكثر من مرحلة انتقالية: معظم الأنواع لا تظهر أي ابتكارات على الإطلاق وقليل منها تظهر الكثير [17].
بالنظر إلى هذا ، يبدو أنه تمدد نقطة للادعاء بأن ما هو في الواقع انقسام في الابتكار هو المسؤول عن التغيير الكمي في حجم الدماغ عبر النظام بأكمله. قد يكون التفسير البديل الواضح هو أن البحث الذكي هو نتاج ثانوي لاكتساب دماغ بحجم أدنى معين (أي اختراق سقف زجاجي لتطور الدماغ).
ما الدليل الذي يشير إلى أن اتخاذ القرار البيئي أو التقني له عواقب فعلية على اللياقة؟ قامت دراسة واحدة فقط بتقييم هذا بشكل مباشر: أظهر ألتمان [67،68] أن إناث قرد البابون التي كانت أكثر قدرة على التوافق مع النظام الغذائي الأمثل (من حيث تناول الطاقة والبروتين) مع بقاء الصغار على قيد الحياة لفترة أطول ، وكان لديهم وظائف إنجابية أطول وأنجبوا نسلًا أكثر ( سواء تم تصنيفها على أنها إجمالي عدد المواليد أو عدد البقاء على قيد الحياة حتى 12 شهرًا).
على الرغم من أن حجم العينة صغير (ن ¼ 6 إناث) ، فإن النتائج خطية ومقنعة بشكل ملحوظ. توفر الأدمغة الكبيرة بالتأكيد القدرة على الانخراط في حل المشكلات بطريقة فعالة عن طريق التجربة والخطأ أو التعلم الثاقب لمحاكمة واحدة [69].
هناك قضية أكثر أهمية تتعلق بافتراض أن الغذاء هو ، أو موازنات الطاقة عن طريق الامتداد ، العامل الأساسي الذي يؤثر على لياقة الحيوان ، إما لأن جميع التأثيرات الخارجية الأخرى تافهة بالمقارنة أو لأن البحث عن الطعام هو العامل الوحيد الذي يمكن للحيوان التحكم به فعليًا من خلال سلوكه .
في الواقع ، بالنسبة للثدييات بشكل عام ، والرئيسيات على وجه الخصوص ، يعتبر الافتراس أيضًا أحد الاعتبارات الرئيسية [70-72] وله تأثير أكبر بكثير على التوزيعات البيوجغرافية للأنواع [19،73] - وهذا على نطاق واسع عبر الثدييات والطيور بشكل عام .
فرديًا وجماعيًا ، يبدو أن الفرضيات الآلية تفشل كتفسير عام لأنها لا توضح سبب احتياج الرئيسيات إلى أدمغة أكبر من أي شخص آخر ، ولماذا تختلف أدمغة الرئيسيات كثيرًا في الحجم بين الأنواع ، ولماذا يجب أن يكون لدى الرئيسيات مجموعات اجتماعية مرتبطة ، ولماذا يجب أن يكون حجم المجموعة يرتبط بحجم الدماغ أو لماذا تحتاج الأنواع المتزاوجة إلى مثل هذه العقول الكبيرة. بشكل عام ، إذن ، قد يتم شرح مهارات البحث عن الطعام بشكل أفضل كحل للقيود المفروضة على نمو الأدمغة الكبيرة والحفاظ عليها عندما تكون مطلوبة لسبب آخر.
يتبع في المشاركة الثانية


Comment